

Preferences in Dogs and Cats
Introduction
Healthy pets exhibit preferences for different nutrient components when provided with foods of similar palatability. Palatability is defined as the subjective gustatory pleasure experienced during food consumption. Research suggests that factors such as volume, quality, energy content, proportions of essential nutrients, and energy density play crucial roles in regulating short-term food intake signals in humans. However, the role of post-ingestive effects on food intake exerts a strong influence on palatability in the long run, often surpassing sensory factors. Ingested foods can trigger physiological changes, and animals learn to associate certain foods with sensory and physiological outcomes through conditioned learning. Animal models can be employed to study the impact of standardized palatability on food intake preferences. Differences in age and body composition may affect animals’ choices of essential nutrients, consequently influencing metabolism. Metabolism can be studied in these animal models based on the circulating concentrations of metabolic intermediates and end products.
Compared to taste, metabolic outcomes related to the intake of essential nutrients might play a more significant role in shaping long-term preferences for fats, carbohydrates, and proteins. Data suggest that there is a clear relationship between the nutrient sensing systems in the oral cavity and the gastrointestinal tract, allowing individuals to appropriately select the quality and quantity of food based on its nutrient composition. The post-ingestive effects of nutrient intake can also modulate food preferences. For example, various sources of fats, especially those high in polyunsaturated fats and low in saturated fats, can influence taste preferences. Certain neuropeptides, such as neuropeptide Y, contribute to integrating food-related information and may directly or indirectly alter dietary patterns, food preferences, fixed dietary habits, and other consequences of nutrient intake.

The feeding behaviors of dogs and cats are typically described with reference to their wild ancestors. Domestication has significantly altered the genotypes of dogs and enhanced their ability to adapt to carbohydrate-rich diets. In domestic cats, the primary genetic changes reported to date are associated with behavioral shifts that occurred when farmers domesticated wild cats to control rodent pests in grain crops, thus losing some of their wild genetic imprints. Cats do not appear to have undergone the same genetic enhancements for carbohydrate metabolism as dogs. Ancestral features of felids can explain why cats prefer higher proportions of protein in their diets compared to dogs. Nevertheless, it is evident that domestic cats can efficiently metabolize most of their calories from carbohydrate-rich foods. Recent experiments suggest that wild carnivorous animals balance essential nutrients by adjusting food intake rather than relying on relatively constant prey availability. Despite this, the diets of wild carnivorous animals are characterized by higher protein and fat content compared to carbohydrates because prey is their primary food source.

Cats are obligate carnivores. Generally, even when allowed to consume tasty foods ad libitum, they can adjust their intake patterns and quantities based on the energy density of the consumed food. Their taste buds are sensitive to amino acids but not to sugars. The relative abundance, novelty, and poor taste of a food can all influence their food choices.
The purpose of this study is to compare the differences in the consumption of essential nutrients in dogs and cats when fed foods with similar palatability. Additionally, the study evaluates the relationship between food intake (nutrient composition) and age, lean body weight or fat body weight, and plasma metabolic profiles. The hypothesis is that when the palatability of food is similar, cats will select different proportions of essential nutrients compared to dogs. Furthermore, it is hypothesized that within a species, choices will vary based on age, lean body weight or fat body weight, leading to unique metabolic profiles in circulating blood that can be used to study the effects of food intake on metabolism.
Research Findings
Palatability Equivalence Testing
Initially, the nutrient content significantly influenced food intake preferences. Dogs favored high-fat foods, while cats preferred high-protein foods.
Food preferences were calculated using consumption ratios, which were determined by dividing the amount of the test food consumed by the total amount of food consumed (value for the grams of the test food/grams of all food consumed). When comparing the consumption of high-fat food (defined as A) to the consumption of other foods (defined as B) in dogs, the A/(A+B) ratio was 0.74 for high-protein food, 0.60 for high-carbohydrate food, and 0.46 for balanced food. The t-test was significant at P<0.05 when B was high-protein or high-carbohydrate food. Subsequently, the quantity of natural chicken flavor enhancer (ranging from 0.5% to 2.5%) was titrated to compensate for the basic changes in nutrient palatability. The least natural chicken flavor enhancer was added to high-fat and balanced foods (0.5%), while the most was added to high-protein food (2.5%). Subsequent calculations comparing the food consumption ratios of high-fat food (A) to other foods (B), with B being high-fat, high-carbohydrate, or balanced food, showed no significant differences (P>0.05).
Since cats initially favored high-protein food, this food was defined as A, and subsequent consumption ratios were calculated to compare the consumption of food A to other foods (defined as B), with B being high-fat, high-carbohydrate, or balanced food. The consumption ratios showed significance at P<0.05 when B was high-fat, high-carbohydrate, or balanced food. Changes in formula ingredients were made to compensate for the basic nutrient palatability. Because cats preferred chicken over pea protein, pea protein (9.0%) and wheat bran (35%) replaced most of the chicken (13.1%) as the protein source in high-protein food. Wheat (20.7%) and brown rice (24.0%) were added in greater amounts than high-protein food (12.0% and 11.4%, respectively) in high-carbohydrate food. Subsequent calculations comparing the food consumption ratios of high-protein food (A) to other foods (B) showed no significant differences (P>0.05).
Nutrient Preferences Study
On average, healthy adult cats select the majority of their calorie intake from carbohydrates (43%) and protein (30%), while healthy adult dogs prefer fats (41%) and carbohydrates (36%) for their calorie intake. The percentages of essential nutrients are based on the comprehensive nutrient intake over a 28-day feeding period (Table 1).
Table 1. Comprehensive Nutrient Intake (Percentage) Over a 28-Day Feeding Period for Dogs and Cats
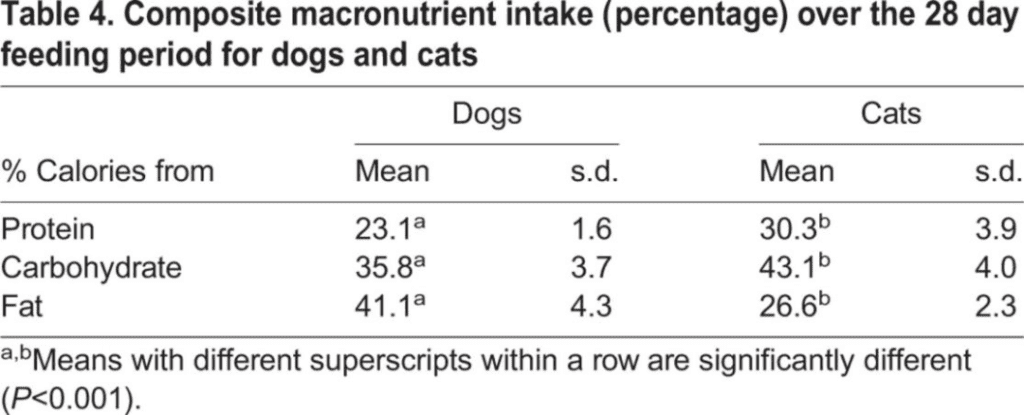
Overall, when fed foods with similar palatability, the intake of essential nutrients by healthy adult dogs and cats differs significantly (P<0.001). Compared to dogs, cats have a broader range of protein intake, as evidenced by the standard deviation in protein intake. None of the dogs or cats selected to maximize their protein calorie intake by exclusively consuming high-protein food.
In cats, an increase in protein calorie intake corresponds to a decrease in carbohydrate calorie intake, as well as a decrease in fat calorie intake. In dogs, changes in protein calorie intake are less pronounced and are not compensated for by carbohydrate calorie intake, although an increase in protein calorie intake results in a decrease in fat calorie intake, similar to cats. Unlike cats, fat and carbohydrate calorie intake in dogs are inversely related; an increase in fat calorie intake corresponds to a decrease in carbohydrate
In general, when feeding foods with similar palatability, healthy adult dogs and cats have different constant nutrient intake levels (P<0.001). Compared to dogs, cats exhibit a wider range of protein intake, as evidenced by the standard deviation of protein intake. Neither dogs nor cats exclusively select food with high protein content to maximize their protein intake.
In cats, as protein intake increases, carbohydrate intake decreases, and fat intake also decreases. In dogs, the variation in protein intake is less pronounced than in cats and is not offset by carbohydrate intake, although increased protein intake is offset by reduced fat intake, similar to cats. Unlike cats, dogs exhibit an inverse relationship between fat and carbohydrate intake; as fat intake increases, carbohydrate intake decreases. In cats, an increase in fat intake does not affect carbohydrate intake.
Age, lean body weight, and fat body weight influence the percentage of calories from protein that dogs and cats choose to consume. In cats, the percentage of calories from protein consumed is negatively correlated with age and lean body weight. Overall, younger cats prefer dietary calories from protein. Cats over 3.2 years of age consume less protein than younger cats. Cats aged 3.2-3.4 years consume the fewest calories from protein. Compared to younger cats with larger lean body weight (>3.8 kg), younger cats with smaller lean body weight prefer to obtain dietary calories from protein (34.4%). In dogs, the percentage of calories from protein chosen for consumption is correlated with age and fat body weight. Overall, dogs with higher fat body weight (>4.2 kg) prefer to obtain dietary calories from protein (24.5%). Age also significantly affects protein intake, with younger dogs with smaller fat body weight (<7.8 years) choosing fewer calories from protein (21.5%).
Based on age and lean body weight (cats), and fat body weight and age (dogs), a regression tree analysis categorizes cats and dogs into groups (four groups for cats, three groups for dogs) to study the effects of constant nutrient intake of selected metabolites in the main constant nutrient categories over 28 days. Plasma DHA concentration is significantly lower (P=0.01) in older cats (>3.4 years) compared to younger cats (<3.2 years) (Table 2). None of the foods contain EPA or DHA components. Plasma phenylalanine concentration is significantly higher (P=0.02) in older cats. Metabolomic analysis of urea cycle intermediates indicates that an increase in protein intake in younger cats is associated with increased concentrations of arginine (P=0.05) and high arginine (P=0.01). Plasma urea concentration does not increase with an increase in protein intake. In older cats, concentrations of sulfated microbial degradation metabolites – para-cresyl sulfate (P=<0.01) and 4-ethylphenyl sulfate (P=0.02) significantly increase.
Table 2. Effects of selected foods consumed over 28 days on selected metabolites in the main constant nutrient categories
Image
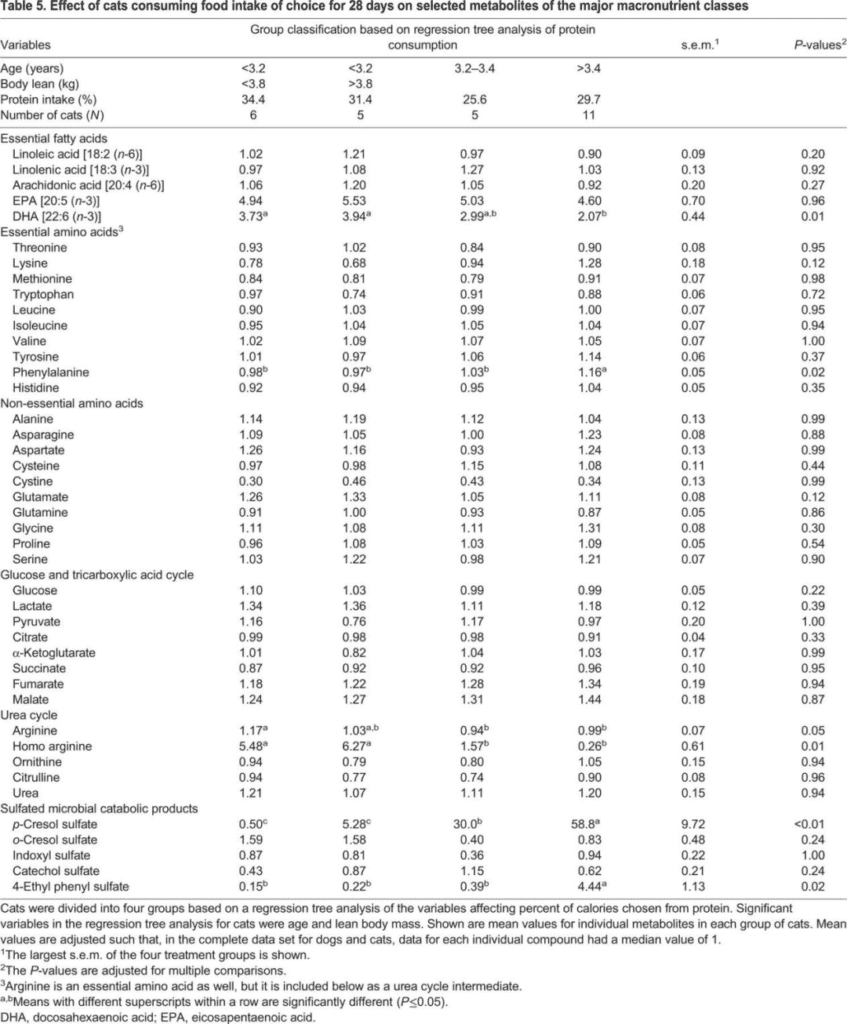
In summary
when feeding foods with similar palatability, the constant nutrient intake levels chosen by healthy adult dogs and cats differ (P<0.0001).
Dogs and cats select specific protein content (average 23% for dogs and 30% for cats) to obtain calories based on factors unrelated to food palatability. Dogs with higher fat body weight prefer dietary calories from protein, while younger dogs with smaller fat body weight consume fewer protein calories.
Overall, younger cats, especially those with smaller lean body weight, prefer dietary calories from protein. Compared to younger lean cats, older cats with moderate protein intake have lower circulating DHA concentrations, higher circulating phenylalanine concentrations, and higher concentrations of sulfated microbial degradation metabolites. Therefore, the circulating metabolite concentrations in cats reflect their food choices. In dogs, there are no differences in selected metabolites in the main constant nutrient categories based on food selection.











